
Page 29 - F. Cell biology
P. 29
MYC suppresses tumor necrosis factor-induced necroptosis by inhibiting the interaction between RIPK1 and
RIPK3
Daehyeon Seong , Manhyung Jeong , Jinho Seo , Eun-Woo Lee , and Jaewhan Song 1
1
2
3
1
1 Department of Biochemistry, College of Life Science and Biotechnology, Yonsei University, Seoul, 120-749, Republic of Korea
2 Environmental Diseases Research Center, Korea Research Institute of Bioscience and Biotechnology (KRIBB), Daejeon 34141, Republic of Korea
3 Metabolic Regulation Research Center, Korea Research Institute of Bioscience and Biotechnology (KRIBB), Daejeon 34141, Republic of Korea
BACKGROUND
MYC is an oncogene that regulates various cellular processes, including proliferation, differentiation, inflammation, and metabolism. We observed that MYC
functions as a negative regulator of necroptosis. Depletion of MYC using siRNA, shRNA, and CRISPR/Cas9 knockout systems accelerated necroptosis, which
was restored to normal levels by reconstitution. Interestingly, both MYC-nick and MYC-NLS mutants, which are predominantly expressed in the cytosol, could
restore necroptosis, suggesting that cytosolic MYC is indispensable as an anti-necroptotic factor. We observed, in detail, that MYC could bind to RIPK3 in the
cytosol blocking the interaction between RIPK1 and RIPK3 upon stimulation of necroptosis. To assess whether this process could be applied to cancer therapy,
we further tested the effect of necroptosis on the suppression of tumorigenesis in a leukemia cell line. The growth of the shMYC-Molm13 cell line skin
xenografts was severely suppressed by the stimulation of necroptosis, implying that necroptosis could suppress cancer cell growth and that its combination
with MYC inhibition may lead to a stronger cancer regression effect.
METHODS
Cell viability analysis and flow cytometry
For the analysis of cell viability, cells were incubated with the CellTiter-Glo reagent and analyzed with a luminometer according to the manufacturer’s protocol (CellTiter-Glo
Luminescent Cell Viability Assay kit, G7571, Promega, Madison, WI, USA). For Annexin V and 7-AAD double staining, prepared cells were harvested and washed with PBS, followed
by incubation with Annexin V-FITC (556547; BD) and 7-AAD (00-6993-50, eBioscience, San Diego, CA, USA) in Annexin V Binding Buffer (51-66121E, BD Biosciences, Franklin Lakes, NJ,
USA) for 15 min according to the manufacturer’s protocol. Dead cells were determined by flow cytometry (BD Accrui C6, BD Biosciences). Data were analyzed using BD Accuri C6
Plus software (BD Biosciences).
Xenograft study
The subcutaneous tumor xenograft model was established by resuspending approximately 5 × 10 Molm13 cells expressing shRNAs in 50 μL of PBS and 50 μL of Matrigel matrix
5
(354234, BD Sciences) and subcutaneously inoculating the mixture into the flank of 6-week-old female Balb/c nude mice (Narabiotech, Seoul, Korea). Six days after inoculation, the
mice bearing xenograft tumors were intraperitoneally injected with birinapant (2 mg/kg) plus emricasan (1 mg/kg). Nineteen days after inoculation, the mice were sacrificed, and
the tumor volumes and masses were measured using a previously described method
RESULTS
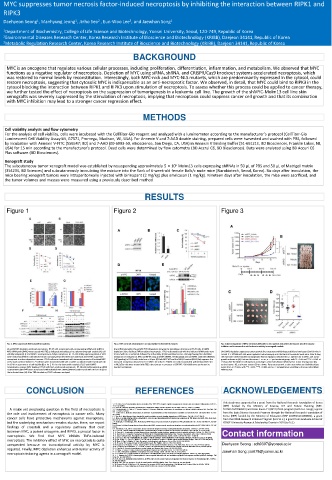
Figure 1 Figure 2 Figure 3
Fig. 1. MYC suppresses TBZ-induced necroptosis. Fig. 2. MYC controls necroptosis in a transcription-independent manner. Fig. 3. Downregulation of MYC sensitizes AML cells to necroptosis induced by birinapant plus the caspase
inhibitor and increases the anti-leukemia activity in xenograft models.
(A and B) MYC depletion facilitates necroptosis. HT-29 cells transfected with a nontargeting siRNA pool (siNT) or (A and B) Immunoblot (A) and qRT-PCR (B) analyses showing the knockdown efficiency in HT-29 cells. (C) MAX
MYC siRNA pool (siMYC) were treated with TBZ as indicated and subjected to a photomicrograph analysis (A), cell depletion fails to facilitate TNFα-mediated necroptosis. HT-29 cells transfected with the indicated siRNA pools were (A-D) MYC depletion suppresses tumor growth after treatment with birinapant (Bir) and emricasan (Emri) in vivo. A
viability analysis (B). (C and D) MYC overexpression delays necroptosis. HT-29 cells stably expressing Mock or MYC treated with TBZ as indicated, followed by cell viability. (D) Immunofluorescence staining showing the subcellular total of 5 × 10 5 Molm13 cells were implanted subcutaneously into the flank of 6-week-old nude mice. After 6 days,
were treated with TBZ for the indicated times and analyzed as described in (A and B). (E and F) MYC suppresses localization of endogenous MYC and RIPK3 using anti-MYC (#5605, Cell Signaling) and anti-RIPK3 antibodies (#13526, the mice were treated with Bir (2 mg/kg) plus Emri (1 mg/kg) as indicated by i.p. injection for 2 weeks, and tumor
necroptosis in a dose-dependent manner. HT-29 cells were transduced with increasing amounts of lentiviral MYC Cell Signaling) in HT-29 cells. Scale bars = 20 μm. (E) Both MYC WT and the NLS1/2 mutant (NLS1/2m) suppress the growth is shown in (A). Data are the means ± s.e.m., n = 7 per treatment group, with *P < 0.05 and ***P < 0.001 at
or the mock control. Stable HT-29 cell lines were transfected with siNT or siMYC as indicated and treated with TBZ increased necroptosis observed in HT-29/MYC KO cells. HT-29/MYC KO cells reconstituted with lentiviral MYC WT 19 days after the Molm13 cell injection according to a two-tailed Mann-Whitney test. Tumor-bearing mice (B),
for 18 h for assessments of cell viability (E) or for 2–4 h for the immunoblot analysis (F). (G and H) MYC or NLS1/2m cells were treated with TBZ in the absence or presence of GSK’963. Cell viability was performed to excised tumors (C), and tumor masses (D) are shown. Data are presented as the mean values and individual data
reconstitution rescues MYC-depleted HT-29 cells from accelerated necroptosis. HT-29 cells stably expressing siRNA- monitor necroptosis. points from n = 7 mice, with **P < 0.01, ***P < 0.001, and n.s. = nonsignificant according to the two-tailed Mann-
resistant MYC (MYC Res#1 ) were transfected with individual MYC siRNA (siMYC#1) and treated with TBZ for 6 h (G) or Whitney test.
the indicated times (H). After TBZ stimulation, HT-29 cells were analyzed.
CONCLUSION REFERENCES ACKNOWLEDGEMENTS
This study was supported by a grant from the National Research Foundation of Korea
1. Y. S. Cho et al., Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation. Cell 137,
1112-1123 (2009). (NRF) funded by the Ministry of Science, ICT and Future Planning (NRF-
A major yet perplexing question in the field of necroptosis is 2. S. He et al., Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha. Cell 137, 1100-1111 (2009). 2015R1A3A2066581) and Brain Korea 21 (BK21) PLUS program (both to J. Song); a grant
3. P. Vandenabeele, L. Galluzzi, T. Vanden Berghe, G. Kroemer, Molecular mechanisms of necroptosis: an ordered cellular explosion. Nat. Rev. Mol. Cell
the role and involvement of necroptosis in cancer cells. Many Biol. 11, 700-714 (2010). from the Basic Science Research Program through the National Research Foundation of
4. L. Galluzzi et al., Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 25, 486-
541 (2018).
Korea (NRF) funded by the Ministry of Education (NRF-2019R1C1C1002831), a grant
cancer cells have protective mechanisms against necroptosis, 5. L. Sun et al., Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase. Cell 148, 213-227 (2012). from the KRIBB Research Initiative Program (to E.W.L.); a grant from Graduate School of
6. J. Zhao et al., Mixed lineage kinase domain-like is a key receptor interacting protein 3 downstream component of TNF-induced necrosis. Proc. Natl. Acad.
Sci. U. S. A. 109, 5322-5327 (2012).
but the underlying mechanism remains elusive. Here, we report 7. T. Delanghe, Y. Dondelinger, M. J. M. Bertrand, RIPK1 Kinase-Dependent Death: A Symphony of Phosphorylation Events. Trends Cell Biol. 30, 189-200 YONSEI UniversityResearch Scholarship Grants in 2020 (to D.S.)
(2020).
findings of crosstalk and a regulatory pathway that exist 8. H. Wang et al., Mixed lineage kinase domain-like protein MLKL causes necrotic membrane disruption upon phosphorylation by RIP3. Mol. Cell 54, 133-146
(2014).
between MYC, a potent oncogene, and RIPK3, a pivotal factor in 9. Y. Dondelinger et al., MLKL compromises plasma membrane integrity by binding to phosphatidylinositol phosphates. Cell Rep 7, 971-981 (2014). Contact information
10. Z. Cai et al., Plasma membrane translocation of trimerized MLKL protein is required for TNF-induced necroptosis. Nat. Cell Biol. 16, 55-65 (2014).
11. X. Chen et al., Translocation of mixed lineage kinase domain-like protein to plasma membrane leads to necrotic cell death. Cell Res. 24, 105-121 (2014).
necroptosis. We find that MYC inhibits TNFα-induced 12. G. Quarato et al., Sequential Engagement of Distinct MLKL Phosphatidylinositol-Binding Sites Executes Necroptosis. Mol. Cell 61, 589-601 (2016).
13. S. Jouan-Lanhouet et al., Necroptosis, in vivo detection in experimental disease models. Semin. Cell Dev. Biol. 35, 2-13 (2014).
necroptosis. The inhibitory effect of MYC on necroptosis is quite 14. J. Seo et al., The roles of ubiquitination in extrinsic cell death pathways and its implications for therapeutics. Biochem. Pharmacol. 162, 21-40 (2019).
15. X. Qin, D. Ma, Y.-x. Tan, H.-y. Wang, Z. Cai, The role of necroptosis in cancer: A double-edged sword? Biochim. Biophys. Acta 1871, 259-266 (2019).
16. B. Shan, H. Pan, A. Najafov, J. Yuan, Necroptosis in development and diseases. Genes Dev. 32, 327-340 (2018).
unexpected because no transcriptional activity by MYC is 17. D. R. Green, The Coming Decade of Cell Death Research: Five Riddles. Cell 177, 1094-1107 (2019). Daehyeon Seong : sdh0307@yonsei.ac.kr
18. A. Degterev, D. Ofengeim, J. Yuan, Targeting RIPK1 for the treatment of human diseases. Proc. Natl. Acad. Sci. U. S. A. 116, 9714-9722 (2019).
19. A. J. Legrand, M. Konstantinou, E. F. Goode, P. Meier, The Diversification of Cell Death and Immunity: Memento Mori. Mol. Cell 76, 232-242 (2019).
required. Finally, MYC depletion enhances anti-tumor activity of 20. Y. Gong et al., The role of necroptosis in cancer biology and therapy. Mol. Cancer 18, 100-100 (2019).
21. S. Philipp, J. Sosna, D. Adam, Cancer and necroptosis: friend or foe? Cell. Mol. Life Sci. 73, 2183-2193 (2016).
necroptosis-inducing agents in a xenograft model. 22. S. Fulda, Therapeutic exploitation of necroptosis for cancer therapy. Semin. Cell Dev. Biol. 35, 51-56 (2014). Jaewhan Song: jso678@yonsei.ac.kr
23. J. R. Cerhan et al., Genetic variation in 1253 immune and inflammation genes and risk of non-Hodgkin lymphoma. Blood 110, 4455-4463 (2007).
24. A. L. Nugues et al., RIP3 is downregulated in human myeloid leukemia cells and modulates apoptosis and caspase-mediated p65/RelA cleavage. Cell
Death Dis. 5, e1384 (2014).
25. G. B. Koo et al., Methylation-dependent loss of RIP3 expression in cancer represses programmed necrosis in response to chemotherapeutics. Cell Res.
25, 707-725 (2015).
26. M. D. Cole, The myc oncogene: its role in transformation and differentiation. Annu. Rev. Genet. 20, 361-384 (1986).
27. M. Conacci-Sorrell, L. McFerrin, R. N. Eisenman, An overview of MYC and its interactome. Cold Spring Harb. Perspect. Med. 4, a014357 (2014).
28. N. Meyer, L. Z. Penn, Reflecting on 25 years with MYC. Nat. Rev. Cancer 8, 976-990 (2008).