
Page 75 - D. Cancer biology
P. 75
Phosphofructokinase 1 Platelet Isoform Promotes β-Catenin Transactivation
for Tumor Development
Jong-Ho Lee 1,2
1 Department of Health Sciences, The Graduate School of Dong-A University
2 Department of Biological Science, Dong-A University, Busan 49315, South Korea
ABSTRACT RESULTS
Metabolism plays a critical role in direct regulation of a
variety of cellular activities via metabolic enzymes and
metabolites. Here, we demonstrate that phosphofructokinase
1 platelet isoform (PFKP), which catalyzes a rate-limiting
reaction in glycolysis, promotes EGFR activation-induced
nuclear translocation and activation of β-catenin, thereby
enhancing the expression of its downstream genes CCND1
and MYC in human glioblastoma cells. Importantly, we
showed that EGFR-phosphorylated PFKP Y64 has a critical
role in AKT activation and AKT-mediated β-catenin S552
phosphorylation and subsequent β-catenin transactivation
and promotion of tumor cell glycolysis, migration, invasion,
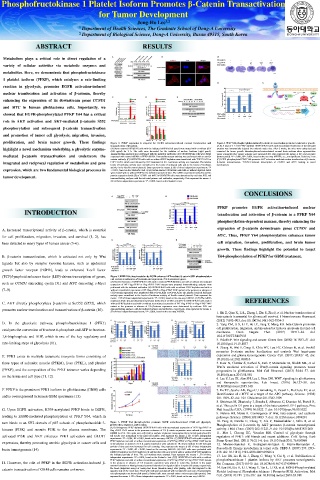
proliferation, and brain tumor growth. These findings Figure 1. PFKP expression is required for EGFR activation-induced nuclear translocation and Figure 4. PFKPY64 phosphorylation-induced β-catenin transactivation promotes brain tumor growth.
transactivation of β-catenin. (A,B) A total of 5 × 105 PFKP-depleted U87/EGFRvIII cells with reconstituted expression of the indicated
highlight a novel mechanism underlying a glycolytic enzyme- (A) Serum-starved U87/EGFR cells with the indicated shRNAs (left panel) were treated with or without EGF proteins was intracranially injected into athymic nude mice. After 2 weeks, the mice were euthanized and
(100 ng/ml) for 9 h. The cells were harvested for the isolation of nuclear fractions (right panel). examined for tumor growth. Hematoxylin-and-eosin–stained coronal brain sections show representative
mediated β-catenin transactivation and underscore the Immunoblotting analyses were performed with the indicated antibodies. (B) U87/EGFRvIII cells were stably tumor xenografts (A). Tumor volumes were measured (B). Data represent the means ± SD of seven mice
(bottom panel). *P < 0.001, #P < 0.001, based on the one-way ANOVA; n.s., not significant. Scale bar, 2 mm.
expressed with control shRNA or PFKP shRNA. Immunofluorescent staining was performed with an anti-β-
catenin antibody. (C) U87/EGFR cells with or without PFKP depletion were transfected with TOP-FLASH or (C) EGFR-phosphorylated PFKP Y64 promotes AKT activation-mediated nuclear translocation of β-catenin,
integrated and reciprocal regulation of metabolism and gene FOP-FLASH, which was followed by EGF treatment for 6 h. Luciferase activity was measured. The relative β-catenin transactivation, TCF/LEF-induced transcription of CCND1 and MYC, leading to tumor
development.
levels of luciferase activity were normalized to the levels of untreated cells and to the levels of luciferase
activity in the Renilla control plasmid. Data represent the means ± SD of three independent experiments. *P
expression, which are two fundamental biological processes in < 0.001, based on the Student's t-test. (D,E) Serum-starved U87/EGFR cells with or without depleted PFKP
were treated with or without EGF for the indicated periods of time. The mRNA expression levels (D) and the
tumor development. protein expression levels (E) of CCND1 and MYC in U87/EGFR cells were determined by real-time PCR and
immunoblotting analyses with the indicated primers and antibodies, respectively. Data represent the means ±
SD of three independent experiments. *P < 0.001, based on the Student's t-test.
CONCLUSIONS
PFKP promotes EGFR activation-induced nuclear
INTRODUCTION translocation and activation of β-catenin in a PFKP Y64
phosphorylation-dependent manner, thereby enhancing the
A. Increased transcriptional activity of β-catenin, which is essential expression of β-catenin downstream genes CCND1 and
for cell proliferation, migration, invasion, and survival (1, 2), has MYC. Thus, PFKP Y64 phosphorylation enhances tumor
been detected in many types of human cancer (3–6). cell migration, invasion, proliferation, and brain tumor
growth. These findings highlight the potential to target
B. β-catenin transactivation, which is activated not only by Wnt Y64-phosphorylation of PFKP for GBM treatment.
ligands but also by receptor tyrosine kinases, such as epidermal
growth factor receptor (EGFR), leads to enhanced T-cell factor
(TCF)/lymphoid enhancer factor (LEF)-driven transcription of genes, Figure 2. PFKP Y64 phosphorylation by EGFR enhances AKT-mediated β-catenin S552 phosphorylation
and nuclear translocation of β-catenin and expression of its downstream genes.
such as CCND1 (encoding cyclin D1) and MYC (encoding c-Myc) (A) Nuclear fractions of U87/EGFRvIII cells with or without PFKP depletion and with or without reconstituted
expression of WT Flag-rPFKP or Flag-rPFKP Y64F mutant were prepared Immunoblotting analyses were
performed with the indicated antibodies. (B) U87/EGFRvIII cells with or without PFKP depletion and with or
(7–9). without reconstituted expression of WT Flag-rPFKP or Flag-rPFKP Y64F mutant in the presence or absence of
CA β-catenin expression were transfected with TOP-FLASH or FOP-FLASH. The relative levels of luciferase
activity were normalized to the levels of luciferase activity of Renilla control plasmid. Data represent the
means ± SD of three independent experiments. *P < 0.001, based on the one-way ANOVA. (C,D) The mRNA
expression levels (C) and the protein expression levels (D) of CCND1 and MYC in U87/EGFRvIII cells with or REFERENCES
C. AKT directly phosphorylates β-catenin at Ser552 (S552), which without PFKP depletion and with or without reconstituted expression of WT Flag-rPFKP or Flag-rPFKP Y64F
mutant in the presence or absence of CA β-catenin expression were determined by real-time PCR and
promotes nuclear translocation and transactivation of β-catenin (10). immunoblotting analyses with the indicated primers and antibodies, respectively. Data represent the means ±
SD of three independent experiments. *P < 0.001, based on the one-way ANOVA. 1. Shi Z, Qian X, Li L, Zhang J, Zhu S, Zhu J, et al. Nuclear translocation of
beta-catenin is essential for glioma cell survival. J Neuroimmune Pharmacol.
(2012) 7:892–903. doi: 10.1007/s11481-012-9354-3
D. In the glycolytic pathway, phosphofructokinase 1 (PFK1) 2. Yang CM, Ji S, Li Y, Fu LY, Jiang T, Meng FD. beta-Catenin promotes
catalyzes the conversion of fructose 6-phosphate and ATP to fructose- cell proliferation, migration, and invasion but induces apoptosis in renal cell
carcinoma. Onco Targets Ther. (2017) 10:711–24. doi:
1,6-bisphosphate and ADP, which is one of the key regulatory and 10.2147/OTT.S117933
3. Polakis P. Wnt signaling and cancer. Genes Dev. (2000) 14:1837–51. doi:
rate-limiting steps of glycolysis (11). 10.1101/gad.14.15.1837
4. Zhang N, Wei P, Gong A, Chiu WT, Lee HT, Colman H, et al. FoxM1
promotes β-catenin nuclear localization and controls Wnt target-gene
E. PFK1 exists in multiple tetrameric isozymic forms consisting of expression and glioma tumorigenesis. Cancer Cell. (2011) 20:427–42. doi:
10.1016/j.ccr.2011.08.016
three types of subunits: muscle (PFKM), liver (PFKL), and platelet 5. Kaur N, Chettiar S, Rathod S, Rath P, Muzumdar D, Shaikh ML, et al.
Wnt3a mediated activation of Wnt/β-catenin signaling promotes tumor
(PFKP), and the composition of the PFK1 tetramer varies depending progression in glioblastoma. Mol Cell Neurosci. (2013) 54:44–57. doi:
on the tissue and cell type (11, 12). 10.1016/j.mcn.2013.01.001
6. Lee Y, Lee JK, Ahn SH, Lee J, Nam DH. WNT signaling in glioblastoma
and therapeutic opportunities. Lab Invest. (2016) 96:137–50. doi:
10.1038/labinvest.2015.140
F. PFKP is the prominent PFK1 isoform in glioblastoma (GBM) cells 7. He TC, Sparks AB, Rago C, Hermeking H, Zawel L, Da Costa LT, et al.
Identification of c-MYC as a target of the APC pathway. Science. (1998)
and is overexpressed in human GBM specimens (13). 281:1509–12. doi: 10.1126/science.281.5382.1509
8. Shtutman M, Zhurinsky J, Simcha I, Albanese C, D'amico M, Pestell R,
et al. The cyclin D1 gene is a target of the beta-catenin/LEF-1 pathway. Proc
G. Upon EGFR activation, K395-acetylated PFKP binds to EGFR, Natl Acad Sci USA. (1999) 96:5522–7. doi: 10.1073/pnas.96.10.5522
9. Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin
leading to EGFR-mediated phosphorylation of PFKP Y64, which in pathways. Science. (2004) 303:1483–7. doi: 10.1126/science.1094291
turn binds to an SH2 domain of p85 subunit of phosphoinositide 3- Figure 3. PFKP Y64 phosphorylation promotes EGFR activation-induced GBM cell glycolysis, 10. Fang D, Hawke D, Zheng Y, Xia Y, Meisenhelder J, Nika H, et al.
proliferation, migration, and invasion. Phosphorylation of β-catenin by AKT promotes β-catenin transcriptional
kinases (PI3K) and recruits PI3K to the plasma membrane. The (A,B) Endogenous PFKP-depleted U87/EGFRvIII cells with reconstituted expression of WT Flag-rPFKP or activity. J Biol Chem. (2007) 282:11221–9. doi: 10.1074/jbc.M611871200
Flag-rPFKP Y64F mutant in the presence or absence of CA β-catenin expression were cultured in no-serum
DMEM for 24 h. The media were collected for analysis of glucose consumption (A) or lactate production (B). 11. Mor I, Cheung EC, Vousden KH. Control of glycolysis through
activated PI3K and AKT enhances PFK1 activation and GLUT1 All results were normalized to the final cell number. Data represent the means ± SD of three independent regulation of PFK1: old friends and recent additions. Cold Spring Harb
experiments. *P < 0.001, #P < 0.001, based on the one-way ANOVA. (C) U87/EGFRvIII cells with or without Symp Quant Biol. (2011) 76:211–6. doi: 10.1101/sqb.2011.76.010868
expression, thereby promoting aerobic glycolysis in cancer cells and PFKP depletion and with or without reconstituted expression of WT Flag-rPFKP or Flag-rPFKP Y64F mutant 12. Moreno-Sanchez R, Rodriguez-Enriquez S, Marin-Hernandez A,
in the presence or absence of CA β-catenin expression were cultured in 1% serum medium for the indicated
periods of time and were harvested for cell counting. Data represent the means ± SD of three independent
brain tumorigenesis (14). experiments. *P < 0.001, #P < 0.001, based on the one-way ANOVA. (D) U251 cells were pretreated with Saavedra E. Energy metabolism in tumor cells. FEBS J. (2007) 274:1393–
DMSO (Control) or 2-DG (10 mM) for 2 h and then the cells were treated with or without EGF (100 ng/ml) for 418. doi: 10.1111/j.1742-4658.2007.05686.x
the indicated periods of time. The cell numbers were counted. Data represent the means ± SD of three 13. Lee JH, Liu R, Li J, Zhang C, Wang Y, Cai Q, et al. Stabilization of
independent experiments. *P < 0.001, based on the Student's t-test. (E,F) U87/EGFRvIII cells with or without
PFKP depletion and with or without reconstituted expression of WT Flag-rPFKP or Flag-rPFKP Y64F mutant phosphofructokinase 1 platelet isoform by AKT promotes tumorigenesis.
H. However, the role of PFKP in the EGFR activation-induced β- in the presence or absence of CA β-catenin expression were placed in serum-free medium in the upper Nat Commun. (2017) 8:949. doi: 10.1038/s41467-017-00906-9
transwell chamber or Matrigel-coated transwell chamber for migration (E) or invasion (F) assays, respectively. 14. Lee JH, Liu R, Li J, Wang Y, Tan L, Li XJ, et al. EGFR-Phosphorylated
catenin transactivation of GBM cells remains unknown. Six hours (migration assay) or twenty-four hours (invasion assay) after plating, cells that migrated to the Platelet Isoform of Phosphofructokinase 1 Promotes PI3K Activation. Mol
opposite side of the insert were fixed with methanol and stained with hematoxylin and eosin. Representative
microphotographs are shown (left panels). Stained cells were counted under a microscope (right panels). Data Cell. (2018) 70:197–210.e197. doi: 10.1016/j.molcel.2018.03.018
represent the means ± SD of three independent experiments. *P < 0.001, based on the one-way ANOVA.