
Page 11 - K. Development and regeneration
P. 11
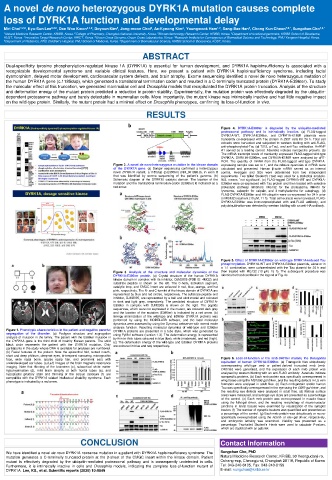
A novel de novo heterozygous DYRK1A mutation causes complete
loss of DYRK1A function and developmental delay
Miri Choi 1,2,# , Kyu-Sun Lee 3,4,# , Dae Woo Kwon 3,4,# , Doyoun Kim 5 , Jong-moon Choi 6 , Ae-Kyeong Kim 3 , Youngwook Ham 1,2 , Sang-Bae Han 2 , Chong Kun Cheon 7,8,* , Sungchan Cho 1,9,*
1 Natural Medicine Research Center, KRIBB, Korea. 2 College of Pharmacy, Chungbuk National University, Korea. 3 Bionanotechnology Research Center, KRIBB, Korea. 4 Department of functional genomics, KRIBB School of Bioscience,
KUST, Korea. 5 Innovative Target Research Center, KRICT, Korea. 6 Green Cross Genome, Green Cross Laboratories, Korea. 7 Research Institute for Convergence of Biomedical Science and Technology, PNU Yangsan Hospital, Korea.
8 Department of Pediatrics, PNU Children’s Hospital, PNU School of Medicine, Korea. 9 Department of Biomolecular Science, KRIBB School of Bioscience, KUST, Korea.
ABSTRACT
Dual-specificity tyrosine phosphorylation-regulated kinase 1A (DYRK1A) is essential for human development, and DYRK1A haploinsufficiency is associated with a
recognizable developmental syndrome and variable clinical features. Here, we present a patient with DYRK1A haploinsufficiency syndrome, including facial
dysmorphism, delayed motor development, cardiovascular system defects, and brain atrophy. Exome sequencing identified a novel de novo heterozygous mutation of
the human DYRK1A gene (c.1185dup), which generated a translational termination codon and resulted in a C-terminally truncated protein (DYRK1A-E396ter). To study
the molecular effect of this truncation, we generated mammalian cell and Drosophila models that recapitulated the DYRK1A protein truncation. Analysis of the structure
and deformation energy of the mutant protein predicted a reduction in protein stability. Experimentally, the mutation protein was effectively degraded by the ubiquitin-
dependent proteasome pathway and barely detectable in mammalian cells. More importantly, the mutant kinase was intrinsically inactive and had little negative impact
on the wild-type protein. Similarly, the mutant protein had a minimal effect on Drosophila phenotypes, confirming its loss-of-function in vivo.
RESULTS
Figure 4. DYRK1A-E396ter is degraded by the ubiquitin-mediated
proteasomal pathway and is intrinsically inactive. (a) FLAG-tagged
DYRK1A-WT, DYRK1A-E396ter, and DYRK1A-K188R plasmids were
transiently co-expressed with Tau protein in 293T cells for 24 h. Total cell
extracts were harvested and subjected to western blotting with anti-FLAG,
anti-phosphorylated-Tau (at T212, p-Tau), and anti-Tau antibodies. HnRNP
A1 served as a loading control. Asterisks indicate nonspecific proteins. (b)
The mRNA transcript levels of transiently expressed FLAG-tagged wild-type
DYRK1A, DYRK1A-E396ter, and DYRK1A-K188R were analyzed by qRT-
PCR. The quantity of mRNA from the FLAG-tagged wild-type DYRK1A-
Figure 2. A novel de novo heterozygous mutation in the kinase domain expressing sample was set to 1, and the relative quantities of mRNA were
of the DYRK1A gene. (a) Sanger sequencing confirmed a heterozygous calculated and presented. Human β-actin mRNA served as an internal
novel DYRK1A variant, c.1185dup (p.E396ter) (NM_001396.3), in exon 8 control. Averages and SDs were determined from two independent
that was identified by exome sequencing of the patient’s genome. (b) experiments. Two-tailed Student’s t-test was used for a statistical analysis.
Schematic diagram of the DYRK1A catalytic domain. The location of the N.S. means ‵not significant’. (c) FLAG-tagged DYRK1A-WT and DYRK1A-
mutation and the translational termination codon (E396ter) is indicated as a E396ter were co-expressed with Tau protein and then treated with selective
red arrow. proteolytic pathway inhibitors: MG132 for the proteasome, NH4Cl for
lysosome, calpeptin for calpain, and 3-methyladenine for autophagy. (d)
FLAG-DYRK1A-E396ter and HA-ubiquitin were co-expressed for 24 h prior
to MG132 treatment (10 μM, 11 h). Total cell extracts were harvested, FLAG-
DYRK1A-E396ter was immunoprecipitated with anti-FLAG antibody, and
polyubiquitination was detected by western blotting with an anti-HA antibody.
Figure 5. Effect of DYRK1A-E396ter on wild-type DYRK1A-induced Tau
phosphorylation. DYRK1A-WT and DYRK1A-E396ter plasmids, alone or in
a 1:1 combination, were co-transfected with the Tau plasmid for 24 h and
Figure 3. Analysis of the structure and molecular dynamics of the then treated with MG132 (10 μM, 15 h). The subsequent procedure was
DYRK1A-E396ter protein. (a) Crystal structure of the human DYRK1A identical to that described in the legend of Fig. 4c.
kinase domain in complex with its inhibitor, DJM2005 (PDB ID: 4MQ2) and
substrate peptide is shown on the left. The C-helix, activation segment,
catalytic loop, and CMGC insert are coloured in red, blue, orange, and hot
pink, respectively. The N- and C-termini of the kinase domain of DYRK1A are
represented by blue and red circles, respectively. The substrate peptide and
inhibitor, DJM2005, are represented by a ball and stick model and coloured
in dark and light grey, respectively. The predicted structure of DYRK1A-
E396ter in complex with DJM2005 is shown on the right. The peptide
sequences, which were not expressed in the mutant, are coloured dark grey
and the location of the mutation (E396ter) is indicated by a red arrow. (b)
Energy minimization of the wild-type and E396ter DYRK1A proteins was
performed by using the MODELLER software, and the local molecular
dynamics were assessed by using the Dynamut webserver with normal mode
analysis function. Resulting molecular dynamics of wild-type and E396ter
Figure 1. Phenotypic characteristics in the patient and negative parental DYRK1A proteins are presented in a tube style, which was generated by
segregation of the disorder. (a) Pedigree structure and segregation using PyMol software (version 1.3). The deformation energy is represented
analysis of mutation in the family. The patient with the E396ter mutation in by thin to thick tubes coloured in blue (low), white (moderate), and red (high).
the DYRK1A gene is the third child of healthy Korean parents. The solid (c) The deformation energy of the wild-type and E396ter DYRK1A proteins
black circle represents the patient with the DYRK1A mutation. Only are coloured in blue and red, respectively.
participants in the study whose DNA is available for analyses are numbered.
(b) Facial features of the patient. Note the epicanthal fold, tented mouth,
short and deep philtrum, deep-set eyes, bi-temporal narrowing, micrognathic
face, wide nasal bone, sparse scalp hair, and prominent ears with Figure 6. Loss-of-function of the mnb-D401ter mutant, the Drosophila
underdeveloped ear lobes. (c,d,e,f) Images of the brain magnetic resonance equivalent of human DYRK1A-E396ter. (a) Transgenic flies ubiquitously
imaging. Note that thinning of the brainstem (c), subcortical white matter overexpressing wild-type mnb or patient-derived truncated mnb (mnb-
hypomyelination (d), mild brain atrophy at both frontal lobes (e), and D401ter) were generated, and the expression of each mnb protein was
hypoplastic pituitary stalk and thinning of the corpus callosum (f) are analysed by western blotting with an anti-FLAG antibody. Asterisks indicate
compatible with the DYRK1A-related intellectual disability syndrome. Each nonspecific proteins. (b) Each mnb protein was specifically overexpressed in
phenotype is indicated by a red arrow. wing tissue using the MS1096-gal driver, and the resulting defects in L5 vein
formation were analysed in adult flies. (c) Each mnb protein and/or human
Tau was specifically overexpressed in the eye using the GMR-gal driver, and
the resulting eye defects were analysed in adult flies. (d) Retinal surface
areas were measured, and average eye sizes are presented as a percentage
of the control. (e) Each mnb protein was overexpressed in muscle tissue
using the Mhc-gal driver, and the resulting morphology of neuromuscular
junctions in larval tissues were examined by visualization of the synaptic
bouton. (f) The number of synaptic boutons was quantified and presented as
a percentage of the control. (g) Each mnb protein was ubiquitously or neuro-
specifically overexpressed using the Actin5- or elav-gal driver, respectively,
and embryonic lethality was examined. Viability was presented as a
percentage. Two-tailed Student’s t-tests were used to calculate P-values,
which are depicted with an asterisk.
CONCLUSION Contact information
We have identified a novel de novo DYRK1A nonsense mutation in a patient with DYRK1A haploinsufficiency syndrome. The Sungchan Cho, PhD
mutation generates a C-terminally truncated protein at the β-sheet of the CMGC insert within the kinase domain. Patient Natural Medicine Research Center, KRIBB, 30 Yeongudanji-ro,
mutant is effectively degraded by the ubiquitin-mediated proteasomal pathway and is consequently undetected in cells. Ochang-eup, Cheongju-si, Chungbuk 28116, Republic of Korea
Furthermore, it is intrinsically inactive in cells and Drosophila models, indicating the complete loss-of-function mutant of Tel: 043-240-6105, Fax: 043-240-6159
DYRK1A. Lee, KS., et al. Scientific reports (2020) 10:9849 E-mail: sungchan@kribb.re.kr