
Page 91 - D. Cancer biology
P. 91
[D-60] Downregulation of TMEM176B protein is promotes Oncogenic-Ras activated
tumor progression.
Hong-Beum Kim
Department of Premedical Course, Chosun University School of Medicine, 309 Pilmun-daero, Dong-gu,
Gwang-ju 61452,Republic of Korea
Abstract The neoplastic transformation by mutant RAS is thought to require remodeling of expression of an entire set of genes. However, the
mechanism underlying Ras-activated tumor metastasis remains unclear. Here we show that oncogenic Ras decreases TMEM176B transcript
production in mouse fibroblast NIH3T3 cells. TMEM176B transcript was undetectable in H460, A549, and H1299 cells exhibiting high Ras
activity, but was relatively abundant in DMS53 cells containing low Ras activity. Furthermore, we demonstrate that TMEM176B transcription
downregulated by oncogenic H-Ras. The ectopic expression of TMEM176B inhibited the epidermal growth factor (EGF)-stimulated ERK1/2
phosphorylation in H-RasV12-transformed NIH3T3 cells. Finally, our results show that TMEM176B inhibits Ras-induced cell proliferation
and tumor formation by oncogenic H-RasV12-transformed NIH3T3 cells. This study identifies the downregulation of TMEM176B as a
potentially important mechanism by which oncogenic Ras-mediated tumor progression.
Result
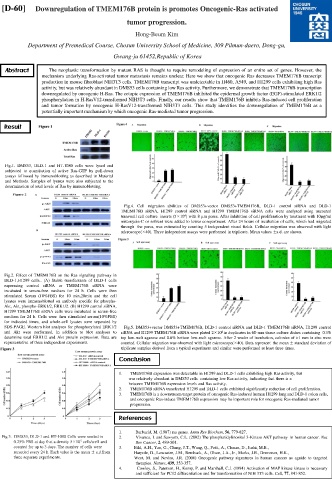
Fig.1. DMS53, DLD-1 and HT-1080 cells were lysed and
subjected to quantitation of active Ras-GTP by pull-down
assays followed by immunoblotting as described in Material
and Methods. Samples of lysates were also subjected to the
determination of total levels of Ras by immunoblotting.
Fig.4. Cell migration abilities of DMS53+vector DMS53+TMEM176B, DLD-1 control siRNA and DLD-1
TMEM176B siRNA, H1299 control siRNA and H1299 TMEM176B siRNA cells were analysed using uncoated
4
transwell cell culture inserts (5×10 ) with 8 µm pores. After inhibition of cell proliferation by treatment with 10µg/ml
mitomycin-C or solvent were added to lower compartment. After 24 hours of incubation of cells, which had migrated
through the pores, was estimated by counting 5 independent visual fields. Cellular migration was observed with light
microscope(×40). Three independent assays were performed in triplicate. Mean values ±s.d. are shown.
Fig.2. Effect of TMEM176B on the Ras signaling pathway in
DLD-1,H1299 cells.. (A) Stable transfectants of DLD-1 cells
expressing control siRNA or TMEM176B siRNA were
incubated in serum-free medium for 24 h. Cells were then
stimulated Serum (10%FBS) for 10 min,20min and the cell
lysates were immunoblotted on antibody specific for phospho-
Akt, Akt, phospho-ERK1/2, ERK1/2. (B) H1299 control siRNA,
H1299 TMEM176B siRNA cells were incubated in serum-free
medium for 24 h. Cells were then stimulated serum(10%FBS)
for indicated times, and whole-cell lysates were separated by
SDS-PAGE. Western blot analyses for phosphorylated ERK1/2 Fig.5. DMS53+vector DMS53+TMEM176B, DLD-1 control siRNA and DLD-1 TMEM176B siRNA, H1299 control
and Akt were performed, in addition to blot analyses to siRNA and H1299 TMEM176B siRNA were plated 2×10 in duplicates in 60-mm tissue culture dishes containing 0.3%
4
determine total ERK1/2 and Akt protein expression. Data are top low-melt agarose and 0.6% bottom low-melt agarose. After 2 weeks of incubation, colonies of >1 mm in size were
representative of three independent experiments. counted. Cellular migration was observed with light microscope(×40). Data represent the mean ± standard deviation of
triplicate samples derived from a typical experiment and similar were performed at least three times.
Conclusion
1. TMEM176B expression was detectable in H1299 and DLD-1 cells exhibiting high Ras activity, but
was relatively abundant in DMS53 cells containing low Ras activity, indicating that there is a
between TMEM176B expression levels and Ras activity.
2. TMEM176B siRNA transfected H1299 and DLD-1 cells exhibited significantly reduction of cell proliferation.
3. TMEM176B is a downstream target protein of oncogenic Ras-induced human H1299 lung and DLD-1 colon cells,
and oncogenic Ras-induce TMEM176B expression may be important role for oncogenic Ras-mediated tumor
progression.
References
1. Barbacid, M. (1987) ras genes. Annu Rev Biochem, 56, 779-827.
Fig.3. DMS53, DLD-1 and HT-1080 Cells were seeded in 2. Vivanco, I. and Sawyers, C.L. (2002) The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat
5
0.25% FBS at day 0 at a density 5×10 cells/well and Rev Cancer, 2, 489-501.
counted for up to 3 days. The number of cells were 3. Bild, A.H., Yao, G., Chang, J.T., Wang, Q., Potti, A., Chasse, D., Joshi, M.B.,
recorded every 24 h. Each value is the mean ± s.d.from Harpole, D., Lancaster, J.M., Berchuck, A., Olson, J.A., Jr., Marks, J.R., Dressman, H.K.,
three separate experiments. West, M. and Nevins, J.R. (2006) Oncogenic pathway signatures in human cancers as aguide to targeted
therapies. Nature, 439, 353-357.
4. Cowley, S., Paterson, H., Kemp, P. and Marshall, C.J. (1994) Activation of MAP kinase kinase is necessary
and sufficient for PC12 differentiation and for transformation of NIH 3T3 cells. Cell, 77, 841-852.