
Page 1 - S. Plant biology
P. 1
AtTPR10 Containing Multiple ANK and TPR Domains Exhibits Chaperone Activity
and Heat-Shock Dependent Structural Switching
Ho Byoung Chae 1 ,† , Su Bin Bae , Seol Ki Paeng , Chang Ho Kang , Joung Hun Park , Eun Seon Lee 1 and Sang Yeol Lee 1,*
1
1
1
1
1 Division of Applied Life Sciences (BK21+) and PMBBRC, Gyeongsang National University, Jinju 52828, Korea.
Abstract
Among the several tetratricopeptide (TPR) repeat-containing proteins encoded by the Arabidopsis thaliana genome, AtTPR10 exhibits an
atypical structure with three TPR domain repeats at the C-terminus in addition to seven ankyrin (ANK) domain repeats at the N-terminus.
However, the function of AtTPR10 remains elusive. Here, we investigated the biochemical function of AtTPR10. Bioinformatic analysis revealed
that AtTPR10 expression is highly enhanced by heat shock compared with the other abiotic stresses, suggesting that AtTPR10 functions as a
molecular chaperone to protect intracellular proteins from thermal stresses. Under the heat shock treatment, the chaperone activity of AtTPR10
increased significantly; this was accompanied by a structural switch from the low molecular weight (LMW) protein to a high molecular weight
(HMW) complex. Analysis of two truncated fragments of AtTPR10 containing the TPR and ANK repeats showed that each domain exhibits a
similar range of chaperone activity (approximately one-third of that of the native protein), suggesting that each domain cooperatively regulates
the chaperone function of AtTPR10. Our results clearly demonstrate that AtTPR10 functions as a molecular chaperone in plants to protect
intracellular targets from heat shock stress.
METHODS
1. Determination of Holdase Chaperone Activity
Holdase chaperone activity was measured using malate dehydrogenase (MDH; Sigma-Aldrich) and citrate synthase (CS) as substrates. MDH
and CS were incubated with AtTPR10, ANK-D, and TPR-D in 50 mM HEPES-KOH (pH 8.0) at 43 °C for 15 min, and the heat-induced thermal
aggregation of both substrates was monitored using a DU800 spectrophotometer (Beckman, Brea, CA, USA) equipped with a thermostatic cell
holder preheated to 43 °C, as described previously
2. Heat Stability Analysis of AtTPR10
To determine the heat stability of AtTPR10, 1 µg each of the recombinant MDH and AtTPR10 proteins was incubated at 23 °C, 50 °C, and 60 °C
for 15 min. Proteins were then centrifuged at 13,000× g for 20 min. The supernatant and pellet fractions were analyzed by sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) using a 12% gel, followed by Coomassie brilliant blue staining. The solubility ratio of
each protein was calculated by dividing the band density of the supernatant fraction with that of the pellet fraction by densitometer.
RESULTS
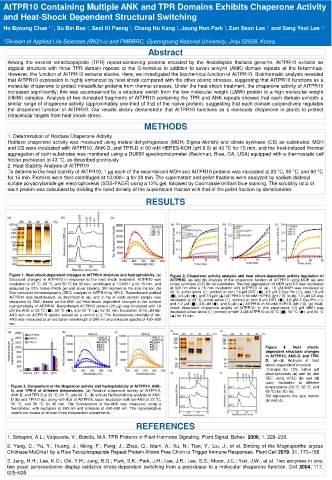
Figure 1. Heat shock-dependent changes in AtTPR10 structure and hydrophobicity. (a) Figure 2. Chaperone activity analysis and heat shock-dependent activity regulation of
Structural changes in AtTPR10 in response to the heat shock treatment. AtTPR10 was AtTPR10. (a) and (b) Analysis of the chaperone function of AtTPR10 using MDH (a) and
incubated at 23 °C, 50 °C, and 60 °C for 30 min, centrifuged at 13,000× g for 15 min, and citrate synthase (CS) (b) as substrates. Thermal aggregation of MDH and CS was monitored
analyzed by 10% native PAGE gel and silver staining. SM represents the size marker. (b) at 340 nm after a 15 min incubation with AtTPR10. In (a), 1.5 µM MDH was incubated at
Size exclusion chromatography (SEC) analysis of AtTPR10 by HPLC. Recombinant purified 43 °C, either alone (○; control) or with 7.5 µM GST (●), 4.5 µM 2-Cys Prx (◇), and 1.5 µM
AtTPR10 was heat-treated, as described in (a), and 2 mg of each protein sample was (■), 4.5 µM (◆), and 7.5 µM (▲) AtTPR10 in 50 mM HEPES (pH 7.0). In (b), 1.2 µM CS was
separated by SEC, based on the MW. (c) Heat-shock dependent changes in the surface incubated at 43 °C, either alone (○; control) or with 6 µM GST (●), 3.6 µM 2-Cys Prx (◇)
hydrophobicity of AtTPR10. Recombinant AtTPR10 protein (20 µg) was incubated with 10 and 1.2 µM (■), 3.6 µM (◆), and 6 µM (▲) AtTPR10 in 50 mM HEPES (pH 7.0). (c) Heat-
shock dependent chaperone activity of AtTPR10. In this experiment, 1.5 µM MDH was
µM bis-ANS at 23 °C (■), 50 °C (◆), and 60 °C (▲) for 20 min. Incubation of 10 µM bis- incubated either alone (○; control) or with 3 µM AtTPR10 at 23 °C (■), 50 °C (◆), and 60 °C
ANS with no AtTPR10 protein served as a control (○). The fluorescence intensity of bis- (▲) for 15 min.
ANS was measured at an excitation wavelength of 390 nm and emission spectra of 430–630
nm.
Figure 4. Heat shock-
dependent structural changes
in AtTPR10, ANK-D, and TPR-
D. (a)–(d) Analysis of heat
shock–dependent structural
changes by 10% native gel
electrophoresis (a) and (c) and
SEC using HPLC (b) and (d)
upon incubation at different
Figure 3. Comparison of the chaperone activity and hydrophobicity of AtTPR10, ANK- temperatures (23 °C, 50 °C, and
D, and TPR-D at different temperatures. (a) Relative chaperone activity of AtTPR10, 60 °C) for 30 min.
ANK-D, and TPR-D at 23 °C, 50 °C, and 60 °C. (b) and (c) Hydrophobicity analysis of ANK- SM represents the size marker
D (b) and TPR-D (c), along with that of AtTPR10, upon incubation with bis-ANS at 23 °C, (a) and (c).
50 °C, and 60 °C for 30 min. The fluorescence of bis-ANS was measured using a
fluorometer, with excitation at 390 nm and emission at 430–630 nm. The representative
results are means of at least three independent experiments.
REFERENCES
1. Schapire, A.L.; Valpuesta, V.; Botella, M.A. TPR Proteins in Plant Hormone Signaling. Plant Signal. Behav. 2006, 1, 229–230.
2. Yang, C.; Yu, Y.; Huang, J.; Meng, F.; Pang, J.; Zhao, Q.; Islam, A.; Xu, N.; Tian, Y.; Liu, J.; et al. Binding of the Magnaporthe oryzae
Chitinase MoChia1 by a Rice Tetratricopeptide Repeat Protein Allows Free Chitin to Trigger Immune Responses. Plant Cell 2019, 31, 172–188.
3. Jang, H.H.; Lee, K.O.; Chi, Y.H.; Jung, B.G.; Park, S.K.; Park, J.H.; Lee, J.R.; Lee, S.S.; Moon, J.C.; Yun, J.W.; et al. Two enzymes in one;
two yeast peroxiredoxins display oxidative stress-dependent switching from a peroxidase to a molecular chaperone function. Cell 2004, 117,
625–635.