
Page 3 - S. Plant biology
P. 3
Chaperone function of Arabidopsis NPR1
Su Bin Bae , Ho Byoung Chae , Seol Ki Paeng , Yong Hun Chi , Seong Dong Wi , Kieu Anh Thi Phan and Sang Yeol Lee 1,*
1
1
1,†
1
1
1
1 The Next-Generation Biogreen Program (SSAC,No.2011),Division of Applied Life Sciences (BK21 Plus), Gyeongsang National University, 501 Jinjudaero,
Jinju52828, Korea
ABSTRACT
Among the defense systems, the NPR1 playing a key role in a plant systemic acquired immune responses has been shown to have multiple functions. The
molecular structure of NPR1 has two domains, BTB/POZ and ANK repeat, that are involved in protein–protein interactions. Despite the function of its SA-
induced defense activity in nucleus, the biochemical property of its cytosolic oligomers has not been elucidated. Based on the results that the reversible
structural change of redox proteins is a typical property of molecular chaperones, we investigated the biochemical characteristics of NPR1. From the study,
the recombinant NPR1 functions as a protein chaperone to protect plants from heat stress through its structural switching by its oligomer form. Under heat-
induced condition, the NPR1 protein prevents from aggregation of substrate. And the structural change was regulated upon the redox changes, such as DTT
treatment dissociated its structure to monomer and reduced its chaperone activity, suggesting that the heat-induced chaperone activity of NPR1 is
dependent on its redox status. In summary, the cytosolic NPR1 oligomer performs the important function of molecular chaperone to protect plants from heat
stress that can be applied to the preparation of heat shock-tolerant useful crops.
METHODS
1. Plasmid cloning and purification of recombinant AtNPR1 in E. coli
AtNPR1 was cloned from an Arabidopsis cDNA library by PCR and AtNPR1 constructs were subcloned into the pET28a binary vector and transformed into
the E. coli BL21 (DE3). Cells were cultured until A600~0.6 and then recombinant 6 × His tagged proteins were cultured with 1 mM isopropyl-β-d-
thiogalactopyranoside (IPTG) for another 12 h at 18 °C. The 6×tagged AtNPR1 proteins underwent dialysis with 50 mM HEPES–KOH (pH 8.0) bufer solution.
Purity of the 6 × His tagged AtNPR1 proteins (2 µg) was tested by SDS-PAGE gel. And different concentrations of DTT were added to protein samples to
check the structural changes of AtNPR1 from oligomer to monomeric forms.
2. Measurement of chaperone activity
The holdase chaperone activity was measured using malate dehydrogenase (MDH, Sigma-Aldrich). Thermal-induced aggregation of MDH was incubated by
additional different concentrations of AtNPR1 in 50 mM HEPES–KOH (pH 8.0) at 43 ℃ for 15 min. Aggregated substrates were monitored using thermo-
controllable DU800 spectrophotometer (Beckman, USA).
RESULTS
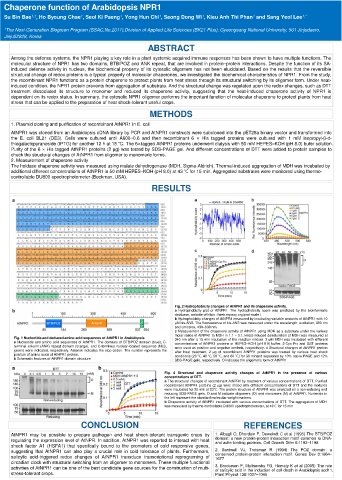
Fig. 2 Hydrophobicity changes of AtNPR1 and its chaperone activity.
a Hydrophobicity plot of AtNPR1. The hydrophobicity score was predicted by the bioinformatic
database, website of https ://web.expasy.org/prot-scale /.
b Hydrophobicity changes of AtNPR1 measured by incubating variable amounts of AtNPR1 with 10
μM bis-ANS. The fluorescence of bis-ANS was measured under the wavelength; excitation, 390 nm
and emission, 430–630 nm.
c Measurement of the chaperone activity of AtNPR1 using MDH as a substrate under the various
molar ratios of AtNPR1 to MDH in 1:1 ~ 3:1. Head-induced denaturation of MDH was measured at
Fig. 1 Nucleotide and deduced amino acid sequences of AtNPR1 in Arabidopsis. 340 nm after a 15 min incubation of the reaction mixture. 2 μM MDH was incubated with different
a Nucleotide and amino acid sequences of AtNPR1. The domains of BTB/POZ domain (blue), C- concentrations of AtNPR1 proteins in HEPES–KOH (pH 8.0) buffer. 2-Cys Prx and GST proteins
terminal ankyrin (ANK) repeat domain (orange), and C-terminus nuclear-located sequence (NLS, were used as positive and negative controls, respectively. d Structural changes of AtNPR1 protein
green) were indicated, respectively. Asterisk indicates the stop codon. The number represents the after heat treatment. 2 μg of recombinant AtNPR1 proteins was treated by various heat shock
position of amino acids of AtNPR1 protein. conditions (23 ℃, 40 ℃, 50 ℃, and 60 ℃) for 30 minand separated by 10% native PAGE and 12%
b Schematic features of AtNPR1 domain structure SDS-PAGE gels, respectively. O indicates the oligomeric form of AtNPR1
Fig. 4 Structural and chaperone activity changes of AtNPR1 in the presence of various
concentrations of DTT.
a The structural changes of recombinant AtNPR1 by treatment of various concentrations of DTT. Purified
recombinant AtNPR1 proteins (2 μg) were mixed with different concentrations of DTT and the mixtures
were incubated for 20 min at 23℃. The protein structure of AtNPR1 was analyzed on a non-reducing and
reducing SDS-PAGE gels. O and M indicate oligomers (O) and monomers (M) of AtNPR1. Numerals in
the left represent the standard molecular weight markers.
b Chaperone activity of AtNPR1 incubated with various concentrations of DTT. The aggregation of MDH
was measured by thermo-controllable DU800 spectrophotometer, at 43℃ for 15 min
CONCLUSION REFERENCES
AtNPR1 may be possible to prepare pathogen and heat shock-tolerant transgenic crops by 1. Albagli O, Dhordain P, Deweindt C et al (1995) The BTB/POZ
regulating the expression level of AtNPR. In addition, AtNPR1 was reported to interact with heat domain: a new protein-protein interaction motif common to DNA-
shock factor A1 (HSFA1) that specifcally bound to the promoters of cold responsive genes, and actin-binding proteins. Cell Growth Difer 6:1193–1198
suggesting that AtNPR1 can also play a crucial role in cold tolerance of plants. Furthermore, 2. Bardwell VJ, Treisman R (1994) The POZ domain: a
salicylic acid-triggered redox changes of AtNPR1 transduce transcriptional reprograming of conserved protein-protein interaction motif. Genes Dev 8:1664–
1677
circadian clock with structural switching from an oligomer to monomers. These multiple functional
activities of AtNPR1 can be one of the best candidate gene sources for the construction of multi- 3. Brodersen P, Malinovsky FG, Hématy K et al (2005) The role
of salicylic acid in the induction of cell death in Arabidopsis acd11.
stress-tolerant crops. Plant Physiol 138:1037–1045