
Page 29 - H. Cell signaling
P. 29
HSV- 1 ICP27 represses NFAT activity through regulating
calmodulin/calcineurin signaling pathway
Mi-jee Kim, Inho Kang, and Jeong Keun Ahn*
Department of Microbiology & Molecular Biology, Chungnam National University, Daejeon, Korea
BACKGROUND AIM
Herpes simplex virus 1 (HSV-1) is a neurotropic human pathogen associated with various symptoms including cold sores, keratitis, blindness, and encephalitis. HSV-1 In this study, we elucidate new mechanism
is able to establish latency in peripheral neurons which can be reactivated for productive infection. During a productive infection in cells, HSV-1 gene expression of immune evasion strategy by HSV-1
proceeded in a tightly regulated cascade. Among the immediate early proteins of HSV-1, ICP27 is a multifunctional protein that is required for viral replication and ICP27. A previous report showed that
viral early gene expression. ICP27 interacts with various cellular proteins and perform multiple functions at both transcriptional and posttranscriptional levels. HSV-1 infection inhibits NFAT activity
Calmodulin (CaM) is a highly conserved calcium binding protein that contains four EF-hand calcium binding motif. CaM acts as a major calcium sensor and relays the and blocks NFAT nuclear translocation.
calcium signaling. Calcineurin is a major CaM binding protein and a ser/thr protein phosphatase controlled by under the control of Ca 2+ /CaM. Calcineurin is involved Therefore, we analyzed the regulatory
in many signaling pathways, such as T-cell activation, the cell cycle, apoptosis, and survival of certain cells. Nuclear factor of activated T cells (NFAT) is a effect of HSV-1 ICP27 on NFAT activity
transcription factor that regulates the expression of many immune response genes. The distinguishing feature of NFAT is its regulation by Ca 2+ /CaM dependent through CaM/calcineurin signaling
phosphatase, calcineurin. When cells are in unstimulated state, inactive phosphorylated form of NFAT localizes in the cytoplasm. An extracellular stimulus increasing pathway.
Ca 2+ signaling leads to calcineurin activation and NFAT dephosphorylation, and translocation of dephosphorylated NFAT.
METHODS
Cells culture HEK 293T cells were maintained in DMEM supplemented with 10% heat-inactivated FBS and 1% penicillin-streptomycin-amphotericin B mixture at 37℃ in humidified atmosphere with 5% CO2. Transfection was performed with
PEI reagent according to the manufacturer’s manual. Plasmids and antibodies The Flag-ICP27 was a gift of Dr. J. Kim (Kim et al., 2008). The vector encoding GFP-NFAT4 was a kind gift from Dr. Kurose. GFP-calcineurin A (CnA) plasmid
was a kind gift from Dr. Gebhardt. ICP27 mutant plasmid was a kind gift from Dr. Sandri-Goldin. Calmodulin 3 gene was cloned into the mammalian expression vector, PEBG to allow it to express GST–CaM. The antibodies used were as follows:
anti-GFP, anti-GST, and anti-Flag. GST pull down assay Cells were transfected with plasmids and incubated for 48hr. Cells were lysed with modified RIPA buffer for 30min at 4℃. The cleared lysate was incubated with glutathione-Sepharose
4B beads for o/n at 4℃. The beads were washed several times with modified RIPA buffer. The pellets were added to SDS sample buffer and boiled for 10min. The samples were resolved on SDS-PAGE for immunoblotting. Western blot analysis
Cell lysates were resolved on SDS-PAGE and the proteins in the gels were transferred onto a PVDF membrane. The membranes were incubated with 5% (w/v) skim milk in PBSt (PBS containing 0.2% Tween 20) and then reacted with primary
antibodies. After washing three times with PBSt, the membranes were incubated with horseradish peroxidase-conjugated anti-IgG. After washing three times with PBSt, the proteins were detected with the ECL reagent (Millipore). Luciferase
assay Luciferase assays was performed using a Dual Luciferase Assay kit (Promega) according to the manufacturer’s instruction. Cells were transiently transfected with luciferase-based reporter constructs (NF-AT-Luc) and pRL-TK Renilla
luciferase plasmid. 48h after transfection, cells were lysed and analyzed with a luminometer (CentroXS LB960, Berthold). Immunofluorescence Cells were splited on sterile galss cover silps and incubated for o/n. 48hr after transfection, cells
were washed with cold PBS and fixed for 10 min with 4% formaldehyde. Cells were washed three times with cold PBS and permeabilized with PBSt for 5min. Cells were incubated with primary antibodies in PBSt (with 0.2% BSA) for o/n and
followed by incubation for 1h with fluorochrome-conjugated secondary antibodies. The fluorescence was examined using a fluorescence microscope. Nuclear fractionation Cells were collected by scraping and lysed by hypotonic lysis buffer.
Cell lysate was passed through a 25G needle 10 times using a 1ml syringe and cell extracts were centrifused at 3000rpm for 5min at 4°C. Remove the supernatant and place in a fresh tube and centrifuge again at 8000 rpm for 20min to cellet
(cytosolic fraction). The nuclear pellet was washed once with fractionation buffer and pass through a 25 G needle 10 times. Centrifuge again at 3000 rpm for 10 min and remove the buffer to collect nuclear fraction.
RESULTS
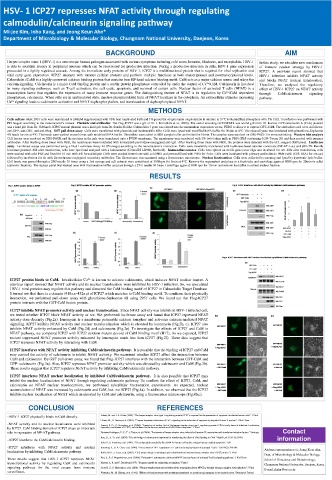
ICP27 protein binds to CaM. Intralcellular Ca 2+ is known to activate calcineurin, which induces NFAT nuclear import. A
previous report showed that NFAT activity and its nuclear translocation were inhibited by HSV-1 infection. So, we speculated
HSV-1 viral proteins may regulate this pathway and dissected the CaM binding motif of ICP27 in Calmodulin Target Database.
It turned out that there is a domain (418a.a~432a.a) of ICP27 which matches to CaM binding motif. To confirme their physically
interaction, we performed pull-down assay with glutathione-Sepharose 4B using 293T cells. We found out that Flag-ICP27
protein interacts with the GST-CaM fusion protein.
ICP27 inhibits NFAT promoter activity and nuclear translocation. Since NFAT activity was inhibits in HSV-1 infected cell,
we tested whether ICP27 block NFAT activity or not. We performed luciferase assay and found that ICP27 repressed NFAT
activity dose-decently (Fig.2a). Ionomycin is a membrane permeable calcium ionophor and activates calcium-mediated NFAT
signaling. ICP27 inhibits NFAT activity and nuclear translocalization which is elevated by ionomycin (Fig.2b, c). ICP27 also
inhibits NFAT activity enhanced by CaM (Fig.2d) and calcineurin (Fig.2e). To investigate the effects of ICP27 and CaM in
NFAT pathway, we compared ICP27 with ICP27 deletion mutant devoid of CaM binding motif (H17). As we expected, ICP27
mutant suppressed NFAT promoter activity enhanced by ionomycin much less than ICP27 (Fig.2f). These data suggest that
ICP27 represses NFAT activity by interacting with CaM.
ICP27 interferes with NFAT activity inhibiting CaM/calcineurin pathway. It is possible that the binding of ICP27 and CaM
may control the activity of calcineurin to inhibit NFAT acitivity. We examined whether ICP27 affect the interaction between
CaM and calcineurin. By GST pull down assay, we found that Flag-ICP27 interferes with the interaction between GST-CaM and
GFP- calcineurin (Fig.3a). Also, ICP27 represses NFAT promoter activity which was elevated by calcineurin and CaM (Fig.3b).
These results suggest that ICP27 regulates NFAT activity by inhibiting CaM/calcineurin pathway.
ICP27 interferes NFAT nuclear localization by inhibited CaM/calcineurin pathway. It is also possible that ICP27 may
inhibit the nuclear localizataion of NFAT through regulating calcineurin pathway. To confirm the effect of ICP27, CaM, and
calcineurin on NFAT nuclear translocatoion, we performed subcellular fractionation experiments. As expected, nuclear
accumulation of NFAT was increased by calcineurin and CaM, but not ICP27 (Fig.4a). In addition, we observed that the ICP27
inhibits nuclear localization of NFAT which an elevated by CaM and calcineurin, using a fluorescence microscope (Fig.4b,c).
CONCLUSION REFERENCES
- HSV-1 ICP27 physically binds to CaM directly. Aubert, M. and J. A. Blaho (1999). "The herpes simplex virus type 1 regulatory protein ICP27 is required for the prevention of apoptosis in infected human cells." J Virol
Chami, M., D. Ferrari, et al. (2003). "Caspase-dependent alterations of Ca2+ signaling in the induction of apoptosis by hepatitis B virus X protein." J Biol Chem
-NFAT activity and its nuclear localization were inhibited Everett, R. D., G. Sourvinos, et al. (2004). "Formation of nuclear foci of the herpes simplex virus type 1 regulatory protein ICP4 at early times of infection: localization,
by ICP27. CaM binding domain of ICP27 plays an important dynamics, recruitment of ICP27, and evidence for the de novo induction of ND10-like complexes." J Virol
role in repression of NF-AT pathway. Fontaine-Rodriguez, E. C., T. J. Taylor, et al. (2004). "Proteomics of herpes simplex virus infected cell protein 27: association with translation initiation factors." Virology Contact
Hou, Q., X. Yi, et al. (2004). "The salt bridge of calcineurinis important for transferring the effect of CNB binding to CNA." FEBS Lett 577(1-2): 294-8. information
- ICP27 interferes the CaM/calcineurin binding.
Itoh, N., S. Yonehara, et al. (1991). "The polypeptideencoded by the cDNA for human cell surface antigen Fas can mediate apoptosis." Cell
-ICP27 interferes with NFAT activity and nuclear Kinoshita, S., B. K. Chen, et al. (1998). "Host control of HIV-1 parasitism in T cells by the nuclear factor of activated T cells." Cell 95(5): 595-604. Address correspondence to: Jeong Keun Ahn,
localization by inhibiting CaM/calcineurin pathway. Koffa, M. D., J. Kean, et al. (2003). "CK2 protein kinase is stimulated and redistributed by functional herpes simplex virus ICP27 protein." J Virol
Dept. of Microbiology & Molecular Biology,
These results suggest that HSV-1 ICP27 represses NFAT Puri, S., B. S. Magenheimer, et al. (2004). "Polycystin-1 activatesthe calcineurin/NFAT (nuclear factor of activated T-cells) signaling pathway." J Biol Chem School of Bioscience and Biotechnology,
transcriptional activity by regulating CaM and calcineurin Rhoads, A. R. and F. Friedberg (1997). "Sequence motifs for calmodulin recognition." Faseb J Chungnam National University, Daejeon, Korea.
signaling pathway for the viral escape from immune Scott, E. S., S. Malcomber, et al. (2001). "Nuclear translocation and activation of the transcriptionfactor NFAT is blocked by herpes simplex virus infection." J Virol E-mail: jkahn@cnu.ac.kr
surveillance. Watanabe, M., M. Kinuya, et al. (1992). "Effects of foetal treatment with methylazoxymethanol on noradrenergicsynapses in rat cerebralcortex." Pharmacol Toxicol